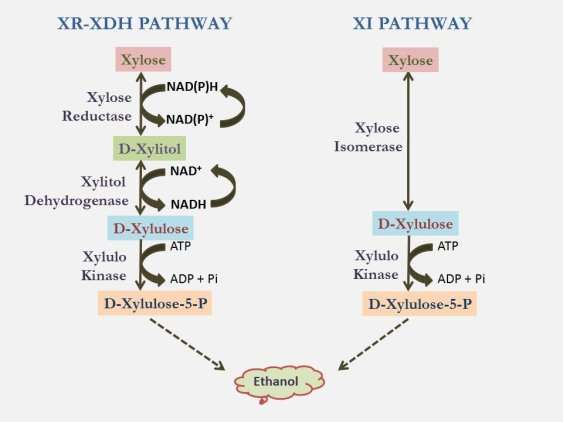
Xylose is the second most abundant sugar in lignocellulosic biomass, comprising up to 35% of the total carbohydrates (Lee, 1997; Saha, 2003; Jeffries and Jin. 2004; Qin et al, 2012). Microorganisms possess three different pathways for D-xylose degradation. The first pathway, present in many bacteria and few fungi, involves xylose isomerase which converts d-xylose to xylulose and phosphorylation of xylulose by xylulokinase followed by its entry to pentose phosphate pathway (Harhangi et al, 2003; Lawlis et al, 1984; Lokman et al, 1991; Rygus et al, 1991, Scheler et al, 1991). The second pathway commonly found in filamentous fungi and yeasts involves use of xylose reductase for converting d-xylose to xylitol, conversion of xylitol to xylulose by xylitol dehydrogenase and conversion of D-Xylulose to D-xylulose 5-phosphate by xylulose kinase followed by its entry into pentose phosphate pathway (Jeffries et al, 1983; Alexander et al, 1988). The third complicated pathway is present in certain Hyperthermophilic Archaea species like Sulfolobus solfataricus and Sulfolobus acidocaldarius. This pathway involves conversion of D-xylose to D-xylonate by xylose Dehydrogenase followed by conversion of D-xylonate to 2-keto-3-xylonate by xylonate dehydratase. 2-keto-3-xylonate is converted to malate (Nunn et al, 2010).
Though many bacteria and filamentous fungi are known xylose utilizers and can be used in fermentation of xylose, yeasts are superior as they yield more ethanol. Among the yeasts Candida shehatae, Pachysolen tannophilus and Pichia stipitis are known to produce ethanol using xylose but require oxygen for xylose utilization (Neirinck et al, 1984; Yu et al, 1995) which reduces the ethanol yield. S.cerevisiae, the yeast used widely for ethanol production cannot metabolize xylose because of the lack of a pathway to efficiently convert xylose to D-xylulose which the organism can metabolize via the pentose phosphate pathway after phosphorylation (Jeffries and Jin. 2004). Genetically engineering S.cerevisiae has been considered to be the viable option for ethanol production from xylose. Though three different pathways exist in microorganisms, the simple pathways involving xylose isomerase, xylose reductase. xylitol dehydrogenase and xylulose kinase are suitable for metabolic engineering of S.cerevisiae as they involve fewer steps. These two pathways have been successfully engineered in S.cerevisiae. Kotter et al (1990) first expressed Xylosereductase and xylitol dehydrogenase genes of Pichia stipites in S.cerevisiae and able to grow S.cerevisiae on xylose as sole carbon source. Expression of P.stipitis xylose reductase in S.cerevisiae using enolase promoter and terminator of Saccharomyces cerevisiae resulted in eylose reductase activity which was 3 time hiher tthan that of P. stipites. Co expression of xylitol dehydrogenase cloned into same plasmid allowed the transformants to grow on a medium containing xylose as the sole carbon source and produce ethanol (Tantirungkij et al, 1993).
Xylose reductase (EC 1.1.1.21)
Xylose reductase (EC 1.1.1.21) is homodimeric oxidoreductase dependent on NADPH or NADH, produced by some yeasts and mycelial fungi which catalyses the reduction of xylose to xylitol (Chiang and Knight, 1960; Kim et al, 1999; Ko et al, 2006). As part of engineering S.cerevisiae for xylose utilization, heterologous xylose reductase genes have been successfully expressed in S.cerevisia (Hallborn et al, 1991; Kim et al, 1999). But the co-factor imbalance between xylose reductase and xylitol dehydrogenase results in xylitol accumulation. Replacement of arginine with histidine (R276H) by point mutation altered the co-factor requirement of xylose reductase from NADPH to NADH (Watanabe et al., 2007) which reduced the xylitol accumulation and enhanced ethanol production. The co-enzyme preference of Candida tenuis xylose reductase towards NADPH was enhanced 170 fold when lysine at position 274 was replaced with arginine and asparagine (ASPN) at position 276 was replaced with ASP (Petschacher et al, 2005; Petschacher and Nidetzky, 2005). This double mutant gene introduced in S.cerevisiae resulted in 42 % enhanced ethanol production compared to the wild type gene containing strain and reduced xylitol accumulation by 52 % (Petschacher and Nidetzky, 2008).
Xylitol dehydrogenase (EC 1.1.1.9):
Xylitol dehydrogenase is a NAD+ dependent enzyme which catalyses the oxidation of xylitol to D-xylulose. Xylitol dehydrogenase genes have been cloned from Galactocandida mastotermitis, Pichia stipites, S.cerviciae, Hypocera jecorina, Aspergillus oryzae and Arxula adeninivorans (Habenicht et al, 1999; Richard et al, 1999; Tran et al, 2004; Kötter et al, 1990). Purification and structural stududies of xylitol dehydrogenase from Galactocandida mastotermitis revealed that xylitol dehydrogenase is a heteronuclear multimetal protein that forms homotetramers and contains 1 mol of Zn2+ ions and 6 mol of Mg2+ ions per mol of 37.4 kDa protomer
Kötter P, Amore R, Hollenberg CP, Ciriacy M (1990) Isolation and characterization of the Pichia stipitis xylitol dehydrogenase gene, XYL2, and construction of a xylose-utilizing Saccharomyces cerevisiae transformant Curr Genet. 18: 493–500.
Tantirungkij M, Nakashima N, Seki T, Yoshida T (1993) Construction of xylose-assimilating Saccharomyces cerevisiae. J Ferment Bioeng 75: 83–88.
Ho NWY, Chen Z, Brainard AP (1998) Genetically engineered Saccharomyces yeast capable of effective cofermentation of glucose and xylose. Appl Environ Microbiol 64: 1852–1859
Walfridsson M, Hallborn J, Penttila M, Keranen S, Hahn-Hägerdal B (1995) Xylose-metabolizing Saccharomyces cerevisiae strains overexpressing the TKL1 and TAL1 genes encoding the pentose phosphate pathway enzymes transketolase and transaldolase. Appl Environ Microbiol 61: 4184–4190.
Toivari MH, Aristidou A, Ruohonen L, Penttilä M (2001) Conversion of xylose to ethanol by recombinant Saccharomyces cerevisiae: Importance of xylulokinase (XKS1) and oxygen availability. Metab Eng 3: 236–249.
Lunzer, R.; Mamnun, Y.; Haltrich, D.; Kulbe, K.D.; Nidetzky, B. Structural and functional properties of a yeast xylitol dehydrogenase, a Zn2+-containing metalloenzyme similar to medium-chain sorbitol dehydrogenases, Biochem. J., 1998, 336, 91-99
NAD+-dependent XDH mutants from P. stipites were developed by introduction of a structural zinc binding site using site-directed mutagenesis (Matsushika et al., 2008).
The differential co-factor requirements of xylose reductase (mainly NADPH) and xylitol dehydrogenase (NAD+) results in co-factor imbalance (redox imbalance) leading to xylitol accumulation (Watanabe et al., 2007). This problem was addressed through protein engineering to synchronize co-factor requirements of xylose reductase and xylitol dehydrogenase (Matsushika et al., 2008; Petschacher and Nidetzky, 2008; Watanabe et al., 2007). Transport has been found to limit the utilization of xylose utilization by wild type yeasts and engineered S.cerevisiae (Eliasson et al, 200; Goffrini et al, 2002; Parachin et al, 2011) and expression of heterologous xylose transporters in S.cerviciae has been found to improve xylose fermentation (Hamacher et al, 2002; Hector et al, 2008; Leandro et al, 2006; Saloheimo et al, 2007).
Xylulokinase (XKS1):
Xylulokinase is an enzyme that converts xylulose to xylulose-5P. S.cervisiae can grown on xylulose, the isomerized form of xylose (Richard et al., 2000; Yang and Jeffries, 1997). The endogenous gene of S.cervisiase (Ho and Chang, 1989) when over expressed significantly enhanced xylose utilization of the Xylose pathway engineered strain (Kotter et al., 1990; Walfridsson et al., 1995; Toivari et al, 2001).
Xylose isomerase:
Xylose isomerase is the enzyme produced by many bacteria and few fungi (Tomoyeda and Horitsu, 1964; Vongsuvanglert and Tani, 1988; Banerjee et al, 1994; Rawat et al, 1996) which catalyses the single step conversion of D-xylose to D-xylulose. Though initial efforts to express bacterial xylose isomerase was unsuccessful, after modifications like codon optimization resulted in expression of heterologous xylose isomerase in S.cerevisiae (Briggs et al, 1984; Sarthy et al, 1987; Amore et al, 1989; Moes et al, 1996; Walfridsson et al, 1996; Gárdonyi et al, 2003; Kuyper et al, 2003; Brat et al, 2009;).
First the D-xylose isomerase gene of anaerobic cellulolytic fungus Piromyces sp. Was expressed in S.cerevisiae but showed poor xylose utilization probably due to the low affinity to D-xylose (Km from 20 to 90 mM) (Kuyper et al, 2003). For example, expression of the Clostridium phytofermentans XI in S. cerevisiae was shown in two separate studies to require codon-optimization and strain adaptation. Expression of xylose isomerase of Clostridium phytofermentans in S.cerevisia needed codon-optimization and strain adaptation (Brat et al, 2009; Ma et al, 2012). Xylose isomerase genes from Ruminococcus flavefaciens XI was recently expressed in S. cerevisiae. Yeast strains expressing several versions of this XI failed to grow aerobically in D-xylose medium, despite one of the expressed XI enzymes having a high specific activity. The XI gene from an anaerobic rumen fungus, Orpinomyces, was expressed in S. cerevisiae (Aeling et al, 2012; Madhavan et al, 2009). Prevotella ruminicola TC2-24 xylose isomerase, when expressed in S. cerevisiae, conferred the ability to grow on D-xylose medium without strain adaption and produced 13.6 g/L ethanol (Hector et al, 2013).
Xylose transporters:
Despite successful metabolic engineering of yeast for xylose utilization transport of xylose, which depends on native glucose transporters, has become a limiting factor for xylose metabolism (Gardonyi et al, 2003; Sedlak et al, 2004; Hector et al, 2008; Leandro et al, 2009). Efforts are underway to engineer existing transporters to alter their specificity towards xylose and introduce heterologous transporters which can efficiently transport xylose into S.cerevisiae (Young et al, 2011; 2014).
GXF1 (glucose/xylose facilitator 1):
GFX1 is the glucose/xylose facilitator (Gxf1) protein mediating high affinity xylose transport in Candida intermedia (Gárdonyi et al, 2003). The gene coding for this protein was isolated (LEANDRO et al, 2006) and expressed in S.cerevisiae. Expression of this gene in a xylose-fermenting laboratory strain of S. cerevisiae increased the rate of xylose utilization and ethanol production at low xylose concentration. (Gárdonyi et al, 2003; Runquist et al, 2009; 2010). Runquist et al, 2010 showed that the GXF1 transporter expressed in S.cerevisae showed very high xylose transport capacity and high growth rate in xylose. The growth was directly proportional to xylose transport kinetics.
Glucose/xylose symporter (GXS):
GXS is a glucose/xylose symporter (GXS) identified from Candida intermedia (Leandro et al., 2006, 2008). Homologue of this gene has been found in Pichia stipitis.
XUT1sugar transporter:
XUT1 is a high affinity sugar transporter isolated from Scheffersomyces stipites which transports xylose. It belongs to the Major Facilitator Superfamily (MFS) (Jeffries et al, 2007).
Xylose permease :
A xylose ABC (ATP-binding cassette) transport operon, xylFGH, was cloned from Thermoanaerobacter ethanolicus , a thermophilic ethanol -producing eubacterium. The cistrons code for a periplasmic D-xylose – binding protein (XylF , partial sequence of 250 amino acids), ATP-binding protein (XylG , 505 amino acids), and integral membrane protein (XylH, 388 amino acids). XylG resembles other eubacterial monosaccharide ABC-ATPases in that its two nucleotide-binding domains (NBDs) are highly homologous, yet significantly different with respect to putative catalytic residues. Unlike most other integral membrane ABC transport proteins, XylH apparently contains 11 or 12 transmembrane segments (TMS) and is similar to a small group of ABC permeases that defy the “2 x 6” helix paradigm. This is the first report of a monosaccharide ABC transport operon in a thermophilic anaerobic eubacterium .
Xylose/proton symporters:
They are active pentose transporters found in xylose-fermenting yeasts (Alcorn and Griffin, 1978; Lucas and van Uden, 1986 ; Kilian and van Uden, 1988; Does and Bisson, 1989; Kilian et al, 1983; Nobre et al,1999) which show high affinity to xylose and specific.
Acetyl xylan esterase:
Acetyl xylan esterase is an enzyme which removes side chains of xylan and this removal enhances xylan hydrolysis (Poutanen et al, 1987). Synergism has been observed between endoxylanase and acetyl xylan esterase improving cellulose hydrolysis (Selig et al, 2008) A direct correlation between removal of acetyl groups and the release of xylose in the hydrolysis of different corn stover substrates with both endoxylanase and AXE was observed (Selig et al, 2009).
Delivering a high-quality product at a reasonable price is not enough anymore.
That’s why we have developed 5 beneficial guarantees that will make your experience with our service enjoyable, easy, and safe.
You have to be 100% sure of the quality of your product to give a money-back guarantee. This describes us perfectly. Make sure that this guarantee is totally transparent.
Read moreEach paper is composed from scratch, according to your instructions. It is then checked by our plagiarism-detection software. There is no gap where plagiarism could squeeze in.
Read moreThanks to our free revisions, there is no way for you to be unsatisfied. We will work on your paper until you are completely happy with the result.
Read moreYour email is safe, as we store it according to international data protection rules. Your bank details are secure, as we use only reliable payment systems.
Read moreBy sending us your money, you buy the service we provide. Check out our terms and conditions if you prefer business talks to be laid out in official language.
Read more