Bioinformatics Analysis of DGAT1 Gene in Domestic Ruminnants
- SIROUS EIDIVANDI
Abstract
Diacylglycerol-O-acyltransferase (DGAT1) gene encodes diacylglyceroltransferase enzyme that playsan important role in glycerol lipid metabolism. DGAT1 is considered to be the key enzyme in controlling the synthesis of triglycerides in adipocytes. This enzyme catalyzes the final step of triglyceride synthesis (transform triacylglycerol (DAG) into triacylglycerol (TAG).
A total of 20 DGAT1 (8,9 Exones) gene sequences belonging to 5 species include cattle (Bos Taurus and BosIndicus), Goats, Sheep and Buffalo were analyzed, and the differentiation within and among the species was also studied. The length of the Exone 8 and Exone 9 respectively were 75bp and 64bp (total: 139bp). Observed genetic diversity was higher among species than within species, and Bos Taurus had more polymorphisms than any other species. Novel amino acid variation sites were detected within several species which might be used to illustrate the functional variation. Differentiation of the DGAT1 gene was obvious among species, and the clustering result was consistent with the taxonomy in the National Center for Biotechnology Information.
Keywords: DGAT1gene, Bioinformatic, Ruminnants
Introduction
Bioinformatics has become an important part of many areas of biology. In experimental molecular biology, bioinformatics techniques such as image and signal processing allow extraction of useful results from large amounts of raw data. In the field of genetics and genomics, it aids in sequencing and annotating genomes and therefore we can observe polymorphic sites, Gene Expressions,Similarities and differences between and within gene sequences in the varies Species and etc. on the other hands gene mapping research has led to the discovery of many polymorphic sites throughout the Ruminants genome that can serve as genetic markers for selection in breeding schemes (Jing-Fen K.et al., 2008).
Diacylglycerolacyltransferases (DGATs) are involved in the process of catalyzation of the final step of the triacylglycerol (TAG) biosynthesis (Hatzopoulos et al., 2011). This enzyme has been found to be encoded by two genes (DGAT1 and DGAT2 ), of which the most studied and important one reavealed to be DGAT1 . This gene is responsible for the codification of the protein related to DGATs activity (Cases et al., 2001).
In bovine, this gene is located on the centromeric end of the bovine chromosome 14 (BTA14), harboring the QTL with a large impact on milk production traits (Grisart et al.,2002; Winter et al., 2002).
DGAT1is a microsomal enzyme catalyzing the addition of fatty acyl Co A to 1, 2, diacylglycerol to yield CoA plus triglycerol and is important in lipogenesis in many tissues, including mammary gland (Kuhnet al.1998). DGAT1 gene is considered to be a very strong positional candidate gene for fat percent of milk.
Kaupeet al., 2004 reported the frequency of this substitution in various cattle breeds and grouped them from very low frequency to fixation inBosindicuscattle breeds. DGAT1 gene is considered to be a very strong positional candidate gene for fat percent of milk. Kaupeet al. [2004] studied Polymorphism of this gene in Bostaurus and Bosindicus breeds. They claimed that K allele of DGAT1 gene is a wild type and the A allele substitution probably occurred after the divergence of Bostaurus and Bosindicus (Kaupeet al., 2004). Recently, many studies showed a significant association between polymorphism of this gene and milk production traits (Grisartet al., 2002; Kharrati Koopaeiet al., 2012; Ripoli MV et al., 2006).
There is a general consensus in the literature that the alanine to lysine amino acid change (K232A) in exon 8 of the DGAT1 gene is associated with reduced milk production (Spelman et al ., 2002; Thaller et al ., 2003a; Banos et al., 2008),
Materials and Methods
A total of 20 sequences with Exons of the DGAT 1 gene and the amino acid sequences belonging to 5 species were obtained from GenBank (Table 1). All the sequences were aligned using the Clustal Omega program implemented in EMBL-EBI service. DnaSP (version 5.1) software was used to analyze the haplotype diversity (Hd), the average number of nucleotide differences (Tajima 1983), the nucleotide diversity (p), synonymous nucleotide diversity (ps), nonsynonymousnucleotide diversity (pa) with the Jukes and Cantor correction, the polymorphic site(S), the singleton variable sites (SP), and the parsimony informative sites (PIP) for each species, and the average number of nucleotide substitutions per site between species (Dxy) (Lynch and Crease, 1990). The phylogenetic tree among 5 species based on the D xy was constructed using the unweight pair group method with the arithmetic mean (UPGMA) implemented in Mega 6 software.
Table 1: DGAT1 gene, Exons 8 and 9 sequences of 5 species
|
Species |
N |
Gene Bank accession number |
|
BosTaurus |
6 |
NM_174693.2 AM263422.1 JQ897353.1 AY065621.1 JQ897351.1 EU077528.1 |
|
Bosindicus |
4 |
DQ435288.1 EU348566.1 EF636701.1 EU348567.1 |
|
Bubalusbubalis |
4 |
JQ627609.1 JF894305.1 NM_001290902 AY999090.1 |
|
Ovisaries |
4 |
EU301803 FJ415875 EU178818 NM_001110164.1 |
|
Capra hircus |
2 |
DQ380249.1 FJ415876.1 |
Results and Discussion
DGAT1 gene, Exon 8,
The Exon sequence of 8 has 75bp in domestic ruminants. We used 6,4,4,4 and 2 sequences of the exon respectively in BosTaurus, Bosindicus, Bubalusbubalis, Ovisaries and Capra hircus(table 1). DnaSP (version 5.1) software was used to analysis of them. The haplotype diversity (Hd) within the sequences of sheep, goat and buffalo was 0, because there wasn’t any polymorphism in these sequences. The haplotype diversity (Hd) within the sequences of bostaurus and bosindicus were shown respectively, 0.733 and 0.5 with 3 and 2 polymorphic sites.
DGAT1 gene, Exon 9,
The Exon sequence of 9 has 64bp in domestic ruminants.The haplotype diversity (Hd) within the sequences of goatbostaurus and bosindicus was 0 but the haplotype diversity (Hd) within the sequences of sheep and Buffalo was 0.5 with 2 polymorphic sites.
Polymorphism and Genetic Diversity among Species
The alignment of 20 sequences of 8 and 9 exons within the region of 139bp and containing gapswas carried out using BioEdit. The results of DnaSP analysis indicated that theselected region (1–140) of the 20 sequences from different species have 139sites, excluding sites with gaps (2). There are 134 invariable (monomorphic) sites and 4 variable (polymorphic) sites that include 3 singleton variable sites and 1 parsimonyinformative sites.The nucleotide diversity (p = 0.00885) and the average number of nucleotide differences ( K = 1.221) for all sequences are lower than the highest values in bostaurus (p = 0.01014 ,K = 1.4).
The polymorphic information and haplotype diversity of the DGAT1 gene (8 and 9Exons) for each species are listed in Table 2.
Table 2: Genetic diversity of the DGAT1gene(8 and 9Exons) in 5 species
|
Species Diversity parameter |
h |
H d |
K |
Π |
πs |
πa |
S |
SP |
PIP |
|
BosTaurus |
3 |
0.733 |
1.4 |
0.01014 |
0 |
0.01459 |
3 |
1 |
2 |
|
Bosindicus |
2 |
0.5 |
1 |
0.00719 |
0.00719 |
– |
2 |
2 |
0 |
|
Bubalusbubalis |
2 |
0.5 |
0.5 |
0.00360 |
0.00360 |
– |
1 |
1 |
0 |
|
Ovisaries |
2 |
0.5 |
0.5 |
0.00360 |
0.00360 |
– |
1 |
1 |
0 |
|
Capra hircus |
1 |
0 |
0 |
0 |
0 |
– |
0 |
0 |
0 |
h, Number of haplotypes; H d, haplotype diversity; K, average number of nucleotide differences; π,
Nucleotide diversityπs, synonymous nucleotide diversity; πa, nonsynonymous nucleotide diversity; S,
Number of polymorphic sites; SP, singleton variable sites; PIP, parsimony informative sites.
The most variable sites (3), singleton variable sites (2), and average number of nucleotide differences (1.4) were found in bostaurus, whichshowed that bostaurus had the highest genetic diversity. Usually, more genetic diversity is most useful for natural selection. The higher genetic diversity of the DGAT1 gene in bostaurus might be related to its extensive adaptability and survival for a polyembryonic animal (Jing-Fen K.et al., 2008).
Amino Acid Variation and Genetic Effects
Higher polymorphism was observed among species than within species, after the 20complete amino acid sequences were aligned using the Clustal Omega program implemented in BioEdit software.
The stop codons in thesequences of the exon 8, 9in Ovisaries, Capra hircus, bosindicus and Bubalus bubalisare onlyUGA but in bos Taurus there are UGA and UAA.Also the exons of bos Taurus had CAC that code histidine and this codone was shown in the other species. The differences between Bos Taurus and the other species in this study maybe related to difference effects of the DGAT 1 gene one the milk production traits. There is a general consensus in the literature that the alanine to lysine amino acid change (K232A) in exon 8 of the DGAT1 gene is associated with reduced milk production (Spelman et al, 2002; Thaller et al, 2003a; Banos et al, 2008),
DNA Divergence and Clustering Analysis
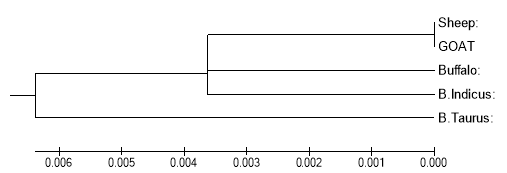
The average number of nucleotide substitutions per site (Dxy) of the DGAT1 gene between species is shown in Table3. Dxyis the index of DNA divergence between or among the sequences. The larger D xy has the smaller the genetic distance. Based on Dxy, a phylogenetic tree was constructed for all the species using the UPGMA method (Fig.1). The divergence time among different species was also labeled on the scale bar calculated from the average nonsynonymous nucleotide rate(0.85 9 10 – 9per year, Li and Dan1991). The dendrogram of different species based on the differentiation of the DGAT1gene agreed with the taxonomy of NCBI. The smallest D xy(0.0000) and divergence showed the closest relationship between Sheep and Goat, which basically accords with that of Yang and Yoder ( 2003) and Wildman et al. (2003). The largest D xy(0.0146) and divergence time displayed the earliest differentiation between BosTaurus and Buffalo, Sheep and Goat, with the average value of 0.0087 for all species(Table 3, Fig1).
Table 3: Average nucleotide substitution per site(Dxy)
|
1 |
2 |
3 |
4 |
5 |
|
|
1. B.Taurus |
|||||
|
2. Buffalo |
0.0146 |
||||
|
3. Sheep |
0.0146 |
0.0072 |
|||
|
4. Goat |
0.0146 |
0.0072 |
0.0000 |
||
|
5. B.Indicus |
0.0073 |
0.0073 |
0.0073 |
0.0073 |
Fig. 1 Phylogenetic tree of the DGAT1(8,9 Exone)gene among 5 species
References
Banos, G., Woolliams, J.A., Woodward, B.W., Forbes,A.B. and Coffey, M.P. (2008) Impact of singlenucleotide polymorphisms in Leptin, LeptinReceptor, Growth Hormone Receptor, andDiacylglycerolAcyltransferase (DGAT1) geneloci on milk production, feed, and body energytraits of UK dairy cows. Journal of Dairy Science91: 3190–3200.
Cases S, Smith SJ, Zheng YW, Myers HM, Lear SR, Sande E, Novak S, Collins C, Welch CB, Lusis AJ,et al.(1998) Identification of a gene encoding an acyl CoA:diacylglycerolacyltransferase, a key enzyme in triacylglycerol synthesis. ProcNatlAcadSci USA 95:13018-13023
Cases S, Stone SJ, Zhou P, Yen E, Tow B, Lardizabal KD, Voelker T, Farese RV (2001). Cloning of DGAT2, a second mammalian diacylglycerolacyltransferase and related family members. Journal of Biological Chemistry 276:38870–38876.
Grisart B, Coppieters W, Farnir F, Karim L, Ford C, Berzi P, Cambisano N, Mni M, Reid S, Simon P, Spelman R, Georges M, Snell R. .( 2002) Positional candidate cloning of a QTL in dairy cattle: Identification of a missense mutation in the bovine DGAT1 gene with major effect on milk yield and composition.Genome Research.12, 222-231.
Grisart B, Farnir F, Karim L, Cambisano N, Kim J, Kvasz A, Mni M, Simori P, Frere J, Coppieters W,et al.(2004) Genetic and functional confirmation of the causality of the DGAT1 K232A quantitative trait nucleotide in affecting milk yield and composition. ProcNatlAcadSci USA 101:2308-2403
Jing-Fen K., Xiang-Long L., Rong-Yan Z., Lan-Hui L., Fu-Jun F. and Xiu-Li G.(2008)Bioinformatics Analysis of Lactoferrin Gene for SeveralSpecies.Biochem Genet 46:312–322
Kaupe B, Winter A, Fries R and Erhardt G (2004) DGAT1 polymorphism inBosIndicusandBostauruscattle breeds. J Dairy Res 71:182-187.
KharratiKoopaei H, Mohammad Abadi MR, Ansari Mahyari S, EsmailizadehKoshkoiyeh A,Tarang AR, Potki P. (2012) Effect of DGAT1 variants on milk composition traits in Iranian Holstein cattlepopulation. Animal Science Papers and Reports. 3, 231-239.
Kuhn CH, Thaller G, Winter A, Bininda-Emonds O, Kaupe B, Erhardt G, Bennewitz J, Schwerin M and Fries R (2004) Evidence for multiple alleles at the DGAT1 locus better explains a quantitative trait locus with major effect on milk fat content in cattle. Genetics 167:1873-1881.
Ripoli MV, Corva P, Giovambattita G. (2006) Analysis of a polymorphism in the DGAT1 gene in 14 cattlebreeds through PCR-SSCP methods. Research Veterinary Science. 80, 287-290
Smith SJ, Cases S, Jensen DR, Chen HC, Sande E, Tow B, Sanan DA, Raber J, Eckel RH and FareseJr RV (2000) Obesity resistance and multiple mechanisms of triglyceride synthesis in mice lacking Dgat. Nat Genet 25:87-90.
Spelman RJ, Ford CA, McElhinney P, Gregory GC and Snell RG (2002) Characterization of the DGAT1 gene in the New Zealand dairy population. J Dairy Sci 85:3514-3517.
Tajima F (1983) Evolutionary relationship of DNA sequences in finite populations. Genetics 105: 437–460
Thaller G, Kuhn C, Winter A, Ewlad G, Bellmann O, Wegner J, Zuhlke H and Fries R (2003) DGAT1, a new positional and functional candidate gene for intramuscular fat deposition in cattle. Anim Genet 34:354-357.
Winter A, Kramer W, Werner F, Kollers S, Kata S, Durstewitz G, Buitkamp J, Womack W, Thaller G and Fries R (2002) Association of a lysine-232/alanine polymorphism in a bovine gene encoding acyl-CoA:diacylglycerolacyltransferase (DGAT1) with variation at a quantitative trait locus for milk fat content. ProcNatlAcadSci USA 99:9300-9305.















